English
English French
French Spanish
Spanish Russian
Russian Korean
Korean Japanese
JapaneseWhat Is Papain and Its Uses in the Feed Industry?
Papain enzymes are primarily found in the roots, stems, leaves, and fruits of papaya, with the highest concentrations in the latex of unripe fruits (Ye Qiteng et al., 1999; Zhao Yuanfan et al., 1999; Yi Yin et al., 2000). Due to their strong protein-degrading ability and the ability to hydrolyze amide bonds and ester bonds, papain enzymes are widely used in the pharmaceutical, food, textile, leather, feed, and dye industries (Wu Xianrong et al., 1988).
1 Overview of Papain Enzymes
Papain derived from papaya juice is not a pure enzyme. These crude enzymes contain not only papain but also lysozyme, cysteine protease, cellulase, glucanase, glutamine, and low-molecular-weight thiol compounds. Based on their isoelectric points, the cysteine-containing enzymes in papaya latex can be classified into three major categories: papain, the complex papain complex (chymopapain), and lysozyme. Papain has a pI of 9.55, chymopapain has a pI of 10.10, and lysozyme has a pI > 11.0. Under near-neutral conditions, these three enzymes can be easily separated by cation exchange column chromatography. In papaya latex, papain accounts for approximately 10% of soluble proteins, chymopapain accounts for 45%, and lysozyme accounts for 20% (Ling Xinghan et al., 1998).

2 Structure of Papain
2.1 Papain
Papain was the first enzyme to be discovered, studied, and widely applied. In 1937, Balls and Lineweaver extracted crystalline papain from fresh papaya juice using a graded salt precipitation method. In 1970, the sequence of papain was determined, and in 1971, its tertiary structure was determined using X-ray crystallography.
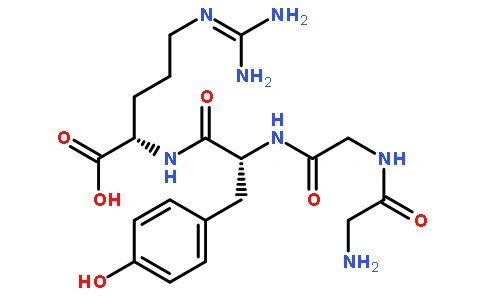
Papain has a relative molecular mass of 21,000 and consists of a single polypeptide chain composed of 212 amino acid residues. In 1969, Husain and Towe reported the amino acid sequence of this enzyme (see Figure 1).
Papain contains seven cysteine residues, six of which form disulfide bonds within three chains, while the remaining free cysteine residue is one of the essential amino acid residues of the active center. Drenth et al. determined the tertiary structure of the papain molecule, which is elliptical in shape and consists of two domains, with a narrow groove at the junction, where the active site of the enzyme is located (Chen Zizhen, 1978).
2.2 Curd Papain
Papain was first discovered, extracted, and named by Jansen and Balls in 1941 (Jansen et al., 1941). The composition of papain is relatively complex. In 1967, Kunimitsu et al. separated papain using ion exchange columns and classified it into two types based on the order of elution: papain A and B; in 1982, Brocklehurst et al. reported that papain consists of four components. In 1985, they used an extremely slow NaCl gradient elution to obtain five active peaks (Kunimitsu et al., 1985; Brocklehurst et al., 1987).
Silvia et al. (1989) conducted circular dichroism analysis on the four subtypes of papain. Currently, it has been discovered that papain exists in different subtypes, with only one or two amino acid differences between them (Buttle et al., 1984). According to data from databases such as EMBL/GenBank/DDJB, there are at least nine subtypes, each encoded by different genes. Among these, six subtypes consist of polypeptide chains with 218 amino acids, two with 227 amino acids, and one with 226 amino acids. The amino acid sequences of the nine subtypes are shown in Figure 2. Comparison results indicate that among these 9 subtypes, only subtype II, subtype III, and subtype V have individual amino acid residues at specific sites that differ from those of other subtypes.
Maes et al. (1996) and Azarkan et al. (1996) determined the three-dimensional structure of papain (see Figure 3). As shown in Figure 3, the polypeptide folding of papain forms two domains of equal size but different shapes—the L domain and the R domain. The L domain is primarily composed of α-helices, while the R domain is primarily composed of anti-parallel β-sheets. The active site is located at the interface between these two domains. Papain belongs to the α+β class, where the C-terminal domain of papain is a fully α-helical structure, while the N-terminal domain is a fully β-sheet structure (Schulz et al., 1979). In contrast, papain belongs to the α/β class, with its secondary structure containing more α-helices and β-sheets, and the folding patterns of the secondary structures are also different (Ssolis-Mendiola et al., 1992).
Papain contains eight cysteine residues, six of which form intra-molecular disulfide bonds, while the remaining two active free sulfhydryl groups are located at residues 25 and 117. Due to their significant distance and different regions, these two residues cannot form intra-molecular disulfide bonds. The two residues have similar activity. From the three-dimensional model of papain, Cys117 is located on the molecular surface, making it easily accessible to derivatization reagents and susceptible to complete and irreversible oxidation. The catalytic Cys25, along with the conserved Cys22 and Cys56, form the S1 microsite. Due to the relatively large “pocket” of the S1 micro-site, it has almost no effect on the enzyme's substrate specificity. However, the S2 micro-site, composed of residues at positions 67, 68, 69, 133, 157, and 207, influences the enzyme's substrate specificity (Maes et al., 1996).
2.3 Papaya lysozyme
In 1955, Smith et al. isolated pure crystalline papain from papaya latex. Later, Haward and Glazer determined the structure and properties of this enzyme. Papain has a molecular weight of 24,000, which is higher than that of animal lysozyme. It consists of 223 amino acids, with the amino acid composition shown in Table 1. Compared to animal lysozyme, papain has significantly higher levels of proline, tyrosine, and phenylalanine.

The arrangement of amino acids at the N-terminal end is: glycine—isoleucine—serine—isoleucine. The arrangement of amino acids at the C-terminal end is: serine—phenylalanine—glycine. The higher structure of papain was determined by circular dichroism spectroscopy, with single helices accounting for approximately 30% of the total. Spectroscopic measurements and chemical reagent reactions indicate that tryptophan residues are entirely embedded within the enzyme molecule and do not participate in the binding of N-acetylglucosamine or active reactions. Papain lysozyme contains 8 cysteine residues, 4 of which form disulfide bonds, and 4 are free SH groups. One SH group is essential for the enzyme's active site. The active site structure of papain lysozyme is significantly different from that of egg white and human lysozyme (Funatsu et al., 1982).
3 Properties of papain
3.1 Properties of papain and papain protease
The structural similarity of papain proteases determines their significant similarity in properties. Both papain and papain protease exhibit broad substrate specificity, Most peptide bonds can be hydrolyzed to some extent by papain, but the rates of hydrolysis vary greatly among different peptide bonds, with some differing by three orders of magnitude; many amino acid derivatives and peptides can serve as substrates, among which arginine derivatives are particularly sensitive to hydrolysis. However, papain cleaves various bonds much faster than papain.
Jansen et al. (1941) reported that papain casein protease hydrolyzes casein at only half the rate of papain. Ebara and Yasunobue used papain casein protease to hydrolyze the β chain of oxidized insulin (oxidized with potassium permanganate), highlighting the enzyme's advantage in hydrolyzing peptide bonds containing acidic amino acid residues and aromatic amino acid residues. When studying the A and B chains of insulin and various peptide segments of different lengths containing glutamic acid, Layle also noted the advantage of papain in hydrolyzing peptide bonds containing glutamic acid. It is worth noting that papain exhibits broader substrate specificity in organic solvent media compared to aqueous media (Jin-Eon So et al., 2000).
Papain and papain chymopapain can also hydrolyze amide bonds and ester bonds (Ling Xinghan et al., 1998). They can catalyze the synthesis of oligopeptides but produce multiple byproducts, which may be due to the broad substrate specificity of the enzyme's S1 site (Jin-Eon So et al., 2000).
The optimal pH values of papain and papain protease vary depending on the substrate. Papain protease, using casein as a substrate, has an optimal reaction temperature of 80°C (pH 7.0) and an optimal pH range of 3–5 at 37°C. The Michaelis constant (Km) value is 1.25 g/l (at pH 7.0 and a reaction temperature of 37°C) (Zucker et al., 1985). The optimal pH value for papain is 7.0, with pH stability ranging from 4 to 9. Enzyme activity remains stable below 60°C and retains some activity up to 90°C. Due to its high thermal stability and good stability, it is suitable for feed processing.
PCMD, chlorobenzyl mercuric acid, iodide acetic acid, iodide acetic acid, hydrogen peroxide, NEM, and heavy metals such as Hg²⁺, Ag⁺, Cu²⁺, and Zn²⁺ can irreversibly inhibit enzyme activity. Papain, containing two free sulfhydryl groups, is more susceptible to oxidation by oxidizing agents or binding with heavy metal ions, leading to inactivation. However, in the presence of various reducing agents (cysteine, thioglycolic acid, glutathione, DTT, etc.) and some metal chelating agents (EDTA), they can reduce the cysteine residues in the active center, thereby activating enzyme activity (Deng Jing et al., 2004).
3.2 Properties of papain lysozyme
Lysozyme is an enzyme that can cleave the β-1,4 bond between N-acetylglucosamine and N-acetylglucosamine. Papain lysozyme is an alkaline protein with an isoelectric point of 10.5, exhibiting only one-third of the activity of egg white lysozyme against Listeria monocytogenes, with an optimal pH of 4.5 and an optimal ionic strength of 0.04–0.07. When decomposing cell walls, it behaves similarly to animal-derived lysozyme, it generates N-acetylmuramic acid at the reducing end. Papain exhibits high activity in decomposing chitin. For example, its activity in decomposing gelatinous chitin is 10 times that of egg white lysozyme, and its activity in decomposing (N-acetylglucosamine)₄ is 200 times that of egg white lysozyme. The products of the enzyme's degradation of (N-acetylglucosamine)₅ are (N-acetylglucosamine)₃ and (N-acetylglucosamine)₂. N-acetylglucosamine does not exist in a free state, and sugar transfer reactions are rarely observed (So et al., 2000).
4 Determination of Enzyme Activity
4.1 Determination of Papain Activity
The activity of papain is determined by measuring the enzyme's ability to catalyze the hydrolysis of substrate proteins under specified conditions (such as a certain temperature and pH value), with the amount of amino acids produced from the hydrolysis of proteins serving as the unit of enzyme activity. The higher the amount of amino acids produced under the same conditions, the greater the enzyme's catalytic reaction capacity and the higher the enzyme activity.
There are three methods for determining papain activity: the UV spectrophotometric method using casein as the substrate, the indophenol colorimetric method, and the BAEE (benzoyl-L-arginine ethyl ester) method. Each method has its own characteristics. The UV spectrophotometric method using casein as a substrate is simple to operate, low-cost, and suitable for comparing enzyme activity during enzyme purification, as well as for determining the activity of papain products and food enzymes. Although the indophenol colorimetric method is cumbersome to perform, it requires simple equipment, making it usable in areas with limited resources; the BAEE method is stable and reproducible, making it suitable for theoretical studies of enzyme properties and the determination of enzyme activity in reagent enzymes (Wu Xianrong, 2005). The method specified in the United States Pharmacopeia and Chinese pharmaceutical standards for determining papain activity is the ultraviolet spectrophotometric method using casein as the substrate (Luo Yanshou, 2000; Chen Deming et al., 2004).
When using the UV spectrophotometric method to detect enzyme activity, the unit of activity is defined as: under the testing conditions, the amount of enzyme required to release the amount of trichloroacetic acid soluble substance from casein hydrolyzed per minute, when the absorbance at a wavelength of 275 nm is equivalent to the absorbance of 1 μg/ml of tyrosine, is one enzyme activity unit.
Several issues require attention when using this method: ① Different trichloroacetic acid concentrations yield different enzyme activity results. ② Casein from different origins exhibits significant differences in papain activity; imported casein has better solubility and higher enzyme activity. ③ Determination must be conducted under optimal pH conditions (Zhao Yuanfan et al., 1999).
4.2 Determination of papain coagulating protease activity
The method for determining the hydrolytic activity of papain coagulating protease uses a UV spectrophotometric method with casein as the substrate. This enzyme also possesses coagulating activity compared to papain, The unit of coagulation activity is defined as the amount of enzyme required to coagulate 1 ml of 10% skim milk (containing 0.01 mol/l CaCl₂) within 40 minutes, which is defined as one enzyme activity unit, i.e., one Soxhlet Unit (SU). The relative activity (RU) is used to indicate the influence of various factors (Arima et al., 1967).
4.3 Papain lysozyme activity determination
There are typically three methods for determining lysozyme activity.
① Using the cell wall as a substrate, the enzyme activity is indicated by the change in turbidity before and after action.
② Using the culture medium of Listeria monocytogenes as a substrate, the enzyme activity is indicated by the change in turbidity before and after action.
Since the above two methods involve solid-liquid phase reactions, it is difficult to accurately measure reaction rates. Therefore, some researchers have prepared a homogeneous substrate using water-soluble polysaccharide hexane diol chitin to determine enzyme activity.
③ Spectrophotometric method: At 450 nm, dissolve a certain amount of freeze-dried bacterial powder or thawed Solubulic bacteria suspension in a certain volume of phosphate-buffered solution to achieve an absorbance value of approximately 1.3. This substrate solution and standard enzyme solution are incubated at 25°C in a water bath. Place 2.5 ml of substrate in a 1 cm cuvette in the water bath, add 0.5 ml of enzyme solution, start timing, record the reading E1 at 1 minute and the reading E2 at 2 minutes, and calculate the enzyme activity using the formula. Different standard enzyme solutions with varying enzyme activities correspond to different ΔE values, and the enzyme activity of the standard enzyme used can be calculated based on these ΔE values, thereby verifying the accuracy of the detection (Zhang Yong, 2004).
5 Immobilization of Papain
Immobilized enzymes are a new technology developed in the 1960s. Immobilization refers to the process of combining free cells or enzymes with a solid insoluble carrier using physical or chemical methods to maintain their activity and enable repeated use. To improve the utilization efficiency of papain and reduce production costs, researchers worldwide have conducted extensive studies on the immobilization and application of papain.
In 1961, J. Cebray successfully immobilized papain on a polyamino acid carrier and used it to hydrolyze fragments of γ-globulin. In 1977, J.W. Finley from the United States employed a glutaraldehyde cross-linking method to immobilize papain on chitin, and used it in the beer production process, achieving good clarification effects. In 1978, P. Monsan et al. in France used aminoalkylated microporous glass as a carrier to cross-link papain on its surface, and also applied it to beer clarification (Ling Xinghan et al., 1998).
Xu Fengcai et al. (1992) used sugarcane bagasse cellulose and nylon as carriers to immobilize papain, determined the enzymatic properties of the immobilized enzyme, and applied it to beer clarification. Li Hong et al. (2001) used chitosan microspheres to immobilize papain, studied the enzymatic properties of the immobilized enzyme, and applied it to casein hydrolysis for tyrosine production. The authors have also conducted a series of studies in this area, including the immobilization of papain using silk fibers from silkworms, investigated the enzymatic properties of the immobilized enzyme and applied it to casein hydrolysis for tyrosine production, as well as silk-immobilized papain in a packed-bed reactor and its application, all of which yielded satisfactory results (Chen Fangyan et al., 2004, 2005a, 2005b), and obtained the corresponding immobilized papain products.

6 Application of papain in the feed industry
During food processing, large amounts of protein by-products are generated, such as animal feathers, animal blood after slaughter, fish trimmings, and fish heads from fish processing. These proteins, when directly ground into feed, are difficult for animals to digest and absorb. Discarding them not only wastes resources but also pollutes the environment. By using proteases to hydrolyze these proteins into soluble small-molecule proteins and amino acids, they become easily digestible and absorbable by animals. This not only enables the development of low-cost, high-quality protein feed but also improves feed utilization efficiency and reduces feed costs (Lü Shimin et al., 2001; Tan Aijuan et al., 1998; Yang Ping et al., 2008). Currently, animal protein enzymes are too expensive for hydrolysis; microbial fermentation methods are prone to contamination by other bacteria and may contain toxins; however, papain has strong activity, high thermal stability, and good stability, making it suitable for feed processing.
Additionally, papain, when added as a feed additive to livestock diets, aids in feed digestion, enhances feed efficiency, reduces feed intake, improves growth rates in livestock, and has significant effects on milk production, milk quality, and the prevention of mastitis in dairy cows.
Zhang Qing et al. (1996) added 0.3% papain to shrimp feed and studied the changes in papain activity during production and its effects in aquaculture. They found that under shrimp feed production conditions (85°C, 45 minutes), papain activity loss was severe. However, under poultry feed production conditions (temperature 75°C), protease activity was preserved quite well. Therefore, under pelletizing conditions, the temperature at which most enzyme activity can be preserved is around 75°C. In shrimp farming, it demonstrated a certain growth-promoting effect, increasing yield by 5%.
Binshiyu et al. (1996) added 0.1% papain to the diet of growing pigs, which improved daily weight gain and feed conversion rate, with the most significant effect observed in pigs weighing 10–20 kg (P < 0.01). The results indicate that adding papain to the diet of growing pigs during the early growth phase is most appropriate, and it can also be used in moderate amounts during the middle growth phase.

He Ting et al. (1992) fed meat-type chicks a diet supplemented with papain (40,000 units/kg of feed). The results showed that although there was no significant difference in average weight gain between the groups supplemented with papain (P > 0.05), feed consumption was slightly lower in all experimental groups compared to the control group. Additionally, the use of FS protein feed (fermented blood meal) in the diet was more effective than fish meal. The reason may be that FS protein feed contains not only animal blood but also various plant fiber-rich cake and meal carriers, which may allow other enzymes in papain to exert their effects.
7 Conclusion
Papain is a pure natural product with strong proteolytic hydrolysis and synthesis capabilities, as well as coagulation, lipolysis, and bacteriolytic activity, making it highly versatile. Currently, the global application ratio of papain is as follows: 75% in the brewing and beverage industry, 10% in the meat processing industry, 5% in the fish processing industry, 3% in the pharmaceutical industry, 5% in the feed processing industry, and 2% in other fields. It is evident that the application ratio of papain in the feed industry is relatively low, indicating significant potential for further development. The reasons may include: the relatively high cost of the enzyme during feed processing, significant loss of enzyme activity, which limits its application; or, as a feed additive, the enzyme is not used scientifically during the feeding process. Therefore, how to more effectively and sustainably enhance and maintain the activity of papain; and how to correctly and reasonably use this enzyme in different animal species and at different stages of feeding to maximize its efficacy are key areas worthy of future research and development efforts.
References
[1]Ye Qiteng, Chen Qiang. Application of Papain [J]. Guangxi Tropical Crops Science and Technology, 1999(4):34-35.
[2]Zhao Yuanfan, Ding Renquan. Processing Technology and Application of Papain [J]. Journal of Yunnan Normal University, 1999, 19(5):46-48.
[3] Yi Yin, Tan Aijuan, Liu Ning. Study on the Production Process of Papain [J]. Guizhou Agricultural Sciences, 2000, 28(5): 24-25.
[4]Wu Xianrong, Zhu Liquan. Papain [J]. Journal of Beijing Agricultural University, 1988, 14(1): 13-17.
[5]Ling Xinghan, Wu Xianrong. Papain and Papaya Cultivation [M]. Beijing: China Agricultural Press, 1998:104.
[6] Chen Zizhen. Food Enzymology [M]. Taiwan: Fuwen Publishing House, 1978: 142.
[7] Kunimitsu K, Yasunoba K T, Chymopapain I V. The chromato - graphic fractionation of partially purified chymopapain and the characterization of crystalline chymopapain [J]. Biochemica et Bio - physica Acta, 1985, 828(2):413-417.
[8]Brocklehurst K, Willenbrock F, Salih E. Cysteine proteinases [J]. New Comprehensive Biochemistry, 1987,16:139-158.
[9]Silvia S M, Rafael Z L, A.Rojo-Dominguez, et al. Structural simi - larity of chymopapain forms as indicated by circular dichroism [J]. Biochemical Journal, 1989, 257:183-186.
[10]Buttle D J, Barrett A J. Chymopapain chromatographic purifica - tion and immunological characterization [J]. Biochemical Journal, 1984, 223(1):81-88.
[11]Maes D, Bouckaert J, Poortmans F, et al. Structure of chymopa - pain at 1.7A resolution[J]. Biochemistry, 1996,35:16292-16298.
[12]Azarkan M, Dominique M, Julie B, et al. Thiol pegylation facili - tates purification of chymopapain leading to diffraction studies at 1.4A resolution [J]. The Journal of Chromatography A, 1996, 749: 69-72.
[13]Schulz G E, Schirmer. Principle of protein structure [M]. New York: SpringerVerlag, 1979:44-107.
[14]Ssolis-Mendiola S, Arroyo-Reyan A, Hernandez-Arana A. Circu - lar dichroism of cysteine proteinases from papaya latex-evidence of differences in the folding of their polypeptide chains.Biochem[J]. Biochemica et Biophysica Acta, 1992, 1118:288-292.
[15] Funatsu, Katsutoshi, and Tsuru, Daisuke. Translated by Li Xingfu. Lysozyme [M]. Jinan: Shandong Science and Technology Press, 1982:74-76.
[16]Jansen E F, Balls A K. Chymopapain: a new crystalline proteinase from papaya latex [J]. The Journal of Biological Chemistry, 1941,137:459-460.
[17]Jin-Eon So, Jong-Shik Shin, Byung-Gee Kim. Protease-catalyzed tripeptide (RGD) synthesis [J]. Enzyme and Microbial Technology, 2000,26:108.
[18]Zucker S, Buttle D J. The proteolytic activities of chymopapain, papain,papaya proteinase Ⅲ [J]. Biochemica et Biophysica Acta, 1985, 828(2):196-204.
[19]Deng Jing, Zhao Shujin. Enzymatic properties of papaya coagulating proteinase [J]. Amino Acids and Biological Resources, 2004, 26(2):18-20.
[20]So J E, Shin J S, Kim B G. Protease catalyzed ripeptide (RGD) synthesis[J]. Enzyme and microbial technology, 2000, 26(2):108 - 114.
[21] Wu, X. R. Development and application of papain [J]. Journal of China Agricultural University, 2005, 10(6):11-15.
[22] Luo, Y. X. Study on the determination method of papain activity [J]. Chinese Journal of Pharmacy, 2000, 35(8):556-558.
[23] Chen Demei, Fu Xiujuan. Applicability of Papain Enzyme Activity Assay Methods in Actual Production [J]. Journal of Guangdong Pharmaceutical University, 2004, 20(3):244-245.
[24] Arima K, Shinjiro I, Gakuzo T. Milk clotting enzyme from microorganisms, Part I, Screening test and identification of potent fungus [J]. Agricultural Biology and Chemistry, 1967, 31(5):540-545.
[25] Zhang Yong. Lysozyme and Its Applications in the Food Industry [J]. Grain Processing and Food Machinery, 2004(3):64-65.
[26] Xu Fengcai, Zhang Wei, Luo Gangyue, et al. Study on the Immobilization of Papain on Sugarcane Bagasse Cellulose and Its Application [J]. Journal of South China Agricultural University, 1992, 13(1):53-59.
[27] Xu Fengcai, Li Mingqi. Study on the Immobilization of Papain on Nylon and Its Application [J]. Biochemistry, 1992, 8(3):302-306.
[28] Li Hong, Wang Weijun, Xu Fengcai. Study on the Characteristics of Chitosan Microsphere-Immobilized Papain [J]. Journal of South China Agricultural University, 2001, 22(2):56-58.
[29] Chen Fangyan, Ji Pingxiong. Study on the Immobilization of Papain on Silkworm Silk Fibroin [J]. Journal of South China Agricultural University, 2004, 25(3):83-86.
[30] Chen Fangyan, Ji Pingxiong. Study on the Characteristics of Papain Immobilized on Silk Fibroin [J]. Journal of South China Agricultural University, 2005a, 26(4):81-83.
[31] Chen Fangyan, Ji Pingxiong. Study on the Filling Bed Reactor of Silk Fibrin-Immobilized Papain and Its Application [J]. Sericulture Science, 2005b, 31(3):286-289.
[32] Lyu Shimin, Tan Aijuan. Hydrolytic action of papain on pig blood meal protein [J]. Guizhou Agricultural Sciences, 2001, 29(4):6-7.
[33] Tan Aijuan, Yang Song, Xiang Langtao. Optimal conditions for papain hydrolysis of feed protein [J]. Journal of Mountain Agriculture and Biology, 1998, 17(2):106-109.
[34] Yang Ping, Xia Yongjun, Fan Weiquan. Hydrolytic effect of papain on tilapia by-products [J]. Fisheries Science, 2008, 27(6):290-292.
[35] Zhang Qing, Li Zhuojiao, Chen Kangle, et al. Study on the addition of papain to shrimp feed [J]. Feed Industry, 1996, 17(5): 8-10.
[36] Bin Shiyu, Pan Shizhong. Application of papain in the diet of growing pigs [J]. Grains and Feed Industry, 1996, 17(7): 24-25.
[37] He Ting, Liang Lin, Pan Suihua, et al. Effects of adding papain to the diet of broiler chicks [J]. Guangdong Journal of Animal Husbandry and Veterinary Science, 1992(2): 9-11.